A new paper at PLoS ONE featuring ancient mitochondrial (mtDNA) data from Wielbark, Przeworsk and early Slavic remains argues for matrilineal continuity in present-day Poland since the Iron Age. It's actually based on a thesis that I blogged about more than two years ago (see here). However, it does include some fresh insights, so it's worth a look even if you read the thesis. RoIA stands for Roman Iron Age.
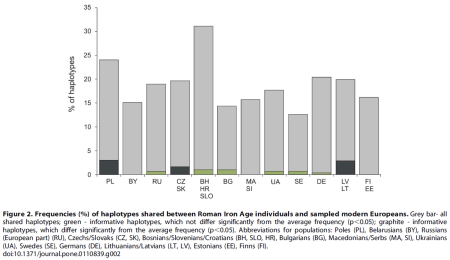
Three modern populations or groups of populations (Lithuanians and Latvians, Poles, and Czechs and Slovaks) were found to contain significantly higher percentages (p,0.05) of shared informative haplotypes with the RoIA samples compared to other present-day populations (Figure 2, Table S4). Notably, modern Poles shared the highest number (nine) of informative mtDNA haplotypes with the RoIA individuals.
...
Of particular interest are three RoIA samples assigned to subhaplogroup H5a1, which were recovered from the Kowalewko (sample K1), the Gaski, and the Rogowo (samples G1 and R3) burial sites (see Figure 1). Recent studies on mtDNA hg H5 have revealed that phylogenetically older subbranches, H5a3, H5a4 and H5e, are observed primarily in modern populations from southern Europe, while the younger ones, including H5a1 that was found among RoIA individuals in our study, date to around 4.000 years ago (kya) and are found predominantly among Slavic populations of Central and East Europe, including contemporary Poles [15]. Notably, we also found one ME sample belonging to subhaplogroup H5a1 (sample OL1 in Table 3). The presence of subclusters of H5a1 in four ancient samples belonging to both the RoIA and the ME periods, and in contemporary Poles, indicates the genetic continuity of this maternal lineage in the territory of modern-day Poland from at least Roman Iron Age i.e., 2 kya.
...
The evolutionary age of H5 sub-branches (,4 kya) [15] also approximates the age of N1a1a2 subclade found in the RoIA population (sample KA2) (Table 2). The coalescence age of N1a1a2 is around 3.4–4 kya, making this haplotype one of the youngest sub-branches within hg N [52]. The N1a1a2 haplotype found in one RoIA individual was classified as unique because no exact match was found among the twelve comparative populations or groups of populations used in the haplotype sharing test. Notably, a similar N1a1a2 haplotype carrying an additional transition at position 16172 was found in a modern-day Polish individual [53].
I suspect the publication of these results at this time, so many months after they were first revealed in the aforementioned thesis, is part of an effort to drum up interest and secure funding for a new project on the genetic history of Greater Poland, which was announced late last year (see here). I say that because one of the people organizing the project, Janusz Piontek, is also listed as a co-author on this paper. So if we're lucky we might soon see full genome sequences from a few of these Iron Age and Medieval samples.
Citation...
Juras A, Dabert M, Kushniarevich A, Malmstro¨m H, Raghavan M, et al. (2014) Ancient DNA Reveals Matrilineal Continuity in Present-Day Poland over the Last Two Millennia. PLoS ONE 9(10): e110839. doi:10.1371/journal.pone.0110839