There's been a lot of horseshit published over the years about Y-chromosome haplogroup R1a, which just happens to be my haplogroup. This includes academic papers in journals like PLoS ONE and Nature.
Indeed, a new paper on the phylogeography of R1a appeared at the Nature website today: Underhill et al. 2014. It's actually a much better effort than anything else on the topic at academic level thus far, but certainly not without issues.
For instance, the authors failed to include two well known and very important R1a subclades in their analysis: the Northwest European-specific R1a-CTS4385 and the East and Central European-specific R1a-Z280. As a result, the former is lumped with R1a-M417* and the latter with R1a-Z282*. In fact, Z280 is shown to be above Z282 in the topology of R1a-M420 (see Figure 1 here), which is plain wrong. These are major oversights and mean that this study is not a very useful resource as far as the phylogeography of European R1a is concerned.
But the paper does show a couple of interesting things. For instance, the maps below offer the best illustration to date of the dichotomy between the European-specific R1a-Z282 and Asian-specific R1a-Z93.
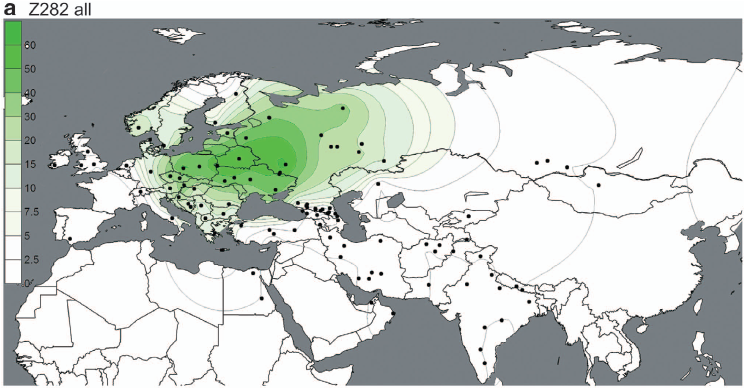

However, these are very closely related subclades, sharing the Z645 mutation (unfortunately not mentioned in the paper), and both reaching high frequencies among Indo-European speakers. It's therefore plausible that groups carrying these markers expanded to the west and east from a zone between their current hotspots, possibly the Volga-Ural region, rather recently.
These migrations had to have happened after 4800-6800 YBP, which is the age of R1a-M417 reported by Underhill et al., and backed up by estimates from genetic genealogists using, among other things, complete R1a sequences (see here). In other words, the rapid expansions of R1a-Z282 and R1a-Z93 appear to have taken place from more or less the same region during the generally accepted early Indo-European timeframe, making them excellent candidates for paternal markers of the early Indo-European dispersals.
At the same time, the paucity of R1a-Z93 and derived lineages in Europe, including Eastern Europe, suggests that historic migrations originating in East and Central Asia, like those of the early Turks, had a negligible effect on the paternal ancestry of modern Europeans. This shows very clearly on the PCA in Figure 4 (see here).
Citation...
Underhill et al., The phylogenetic and geographic structure of Y-chromosome haplogroup R1a, European Journal of Human Genetics, advance online publication, 26 March 2014; doi:10.1038/ejhg.2014.50
See also...
The beast among Y-haplogroups